The ostrich shares the order Struthioniformes with the kiwis, emus, rheas and cassowaries. However, phylogenetic studies have shown that it is the basal extant member of Palaeognathae and is thus equally closely related to flighted tinamous.[3][4] It is distinctive in its appearance, with a long neck and legs, and can run at up to about 70 km/h (20 m/s; 40 mph),[5] the fastest land speed of any bird.[6] The ostrich is the largest living species of bird and lays the largest eggs of any living bird (extinct elephant birds of Madagascar and the giant moa of New Zealand laid larger eggs).
The ostrich's diet consists mainly of plant matter, though it also eats invertebrates. It lives in nomadic groups of 5 to 50 birds. When threatened, the ostrich will either hide itself by lying flat against the ground, or run away. If cornered, it can attack with a kick of its powerful legs. Mating patterns differ by geographical region, but territorial males fight for a harem of two to seven females.
The ostrich is farmed around the world, particularly for its feathers, which are decorative and are also used as feather dusters. Its skin is used for leather products and its meat is marketed commercially.[5]

Description
Ostriches usually weigh from 63 to 145 kilograms (139–320 lb),[5][7] Ostriches of the East African race (S. c. massaicus) averaged 115 kg (254 lb) in males and 100 kg (220 lb) in females, while the nominate subspecies (S. c. camelus) was found to average 111 kg (245 lb) in unsexed adults.[5] Exceptional male ostriches (in the nominate subspecies) can weigh up to 156.8 kg (346 lb).[5] At sexual maturity (two to four years), male ostriches can be from 2.1 to 2.8 m (6 ft 11 in to 9 ft 2 in) in height, while female ostriches range from 1.7 to 2.0 m (5 ft 7 in to 6 ft 7 in) tall.[5] New chicks are fawn in colour, with dark brown spots.[8] During the first year of life, chicks grow at about 25 cm (9.8 in) per month. At one year of age, ostriches weigh approximately 45 kilograms (99 lb). Their lifespan is up to 40–45 years.The feathers of adult males are mostly black, with white primaries and a white tail. However, the tail of one subspecies is buff. Females and young males are greyish-brown and white. The head and neck of both male and female ostriches is nearly bare, with a thin layer of down.[7][8] The skin of the female's neck and thighs is pinkish gray,[8] while the male's is blue-gray, gray or pink dependent on subspecies.
-
 Head
Head
-
 Beak open
Beak open
-
 Ostrich skull
Ostrich skull
-
 Foot
Foot
-
 Claws on the wings
Claws on the wings





Their skin varies in colour depending on the subspecies, with some having light or dark gray skin and others having pinkish or even reddish skin.[5] The strong legs of the ostrich are unfeathered and show bare skin, with the tarsus (the lowest upright part of the leg) being covered in scales: red in the male, black in the female.[5] The tarsus of the ostrich is the largest of any living bird, measuring 39 to 53 cm (15 to 21 in) in length.[5] The bird has just two toes on each foot (most birds have four), with the nail on the larger, inner toe resembling a hoof. The outer toe has no nail.[citation needed] The reduced number of toes is an adaptation that appears to aid in running, useful for getting away from predators. Ostriches can run at a speed over 70 km/h (43 mph) and can cover 3 to 5 m (9.8 to 16.4 ft) in a single stride.[12] The wings reach a span of about 2 metres (6 ft 7 in), and the wing chord measurement of 90 cm (35 in) is around the same size as for the largest flying birds.[5][13] The wings are used in mating displays and to shade chicks. The feathers lack the tiny hooks that lock together the smooth external feathers of flying birds, and so are soft and fluffy and serve as insulation. Ostriches can tolerate a wide range of temperatures. In much of their habitat, temperatures vary as much as 40 °C (100 °F) between night and day. Their temperature control mechanism relies on action by the bird, which uses its wings to cover the naked skin of the upper legs and flanks to conserve heat, or leaves these areas bare to release heat. They have 50–60 tail feathers, and their wings have 16 primary, four alular and 20–23 secondary feathers.[5]
The ostrich's sternum is flat, lacking the keel to which wing muscles attach in flying birds.[14] The beak is flat and broad, with a rounded tip.[7] Like all ratites, the ostrich has no crop,[15] and it also lacks a gallbladder.[16] They have three stomachs, and the caecum is 71 cm (28 in) long. Unlike all other living birds, the ostrich secretes urine separately from faeces.[17] All other birds store the urine and faeces combined in the coprodeum, but the ostrich stores the faeces in the terminal rectum.[17] They also have unique pubic bones that are fused to hold their gut. Unlike most birds, the males have a copulatory organ, which is retractable and 20 cm (8 in) long. Their palate differs from other ratites in that the sphenoid and palatal bones are unconnected.[5]
Taxonomy
The ostrich was originally described by Linnaeus in his 18th-century work, Systema Naturae under its current binomial name.[18] Its scientific name is derived from Latin, struthio meaning "ostrich" and camelus meaning "camel", alluding to its dry habitat.[19]The ostrich belongs to the ratite order Struthioniformes. Other members include rheas, emus, cassowaries, moa, kiwi and the largest bird ever, the now-extinct elephant bird (Aepyornis). However, the classification of the ratites as a single order has always been questioned, with the alternative classification restricting the Struthioniformes to the ostrich lineage and elevating the other groups.
Subspecies

- Common ostrich (S. camelus) complex:
- S. c. australis, Southern ostrich, southern Africa. It is found south of the rivers Zambezi and Cunene. It is farmed for its meat, leather and feathers in the Little Karoo area of Cape Province.[20]
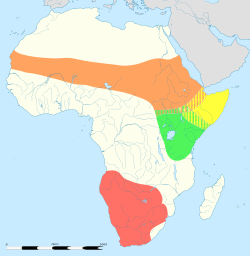
- S. c. camelus, North African ostrich, or red-necked ostrich, North Africa. Historically it was the most widespread subspecies, ranging from Ethiopia and Sudan in the east throughout the Sahel[21] to Senegal and Mauritania in the west, and north to Egypt and southern Morocco, respectively. It has now disappeared from large parts of this range,[22] and it only remains in 6 of the 18 countries where it originally occurred, leading some to consider it Critically Endangered.[23] It is the largest subspecies, at 2.74 m (9.0 ft) in height and up to 154 kilograms (340 lb) in weight.[24] The neck is pinkish-red, the plumage of males is black and white, and the plumage of females is grey.[24]
- S. c. massaicus, Masai ostrich, East Africa. It has some small feathers on its head, and its neck and thighs are pink. During the mating season, the male's neck and thighs become brighter. Its range is essentially limited to southern Kenya and eastern Tanzania[21] and Ethiopia and parts of southern Somalia.[24]
- S. c. syriacus, Arabian ostrich or Middle Eastern ostrich, Middle East. Was formerly very common in the Arabian Peninsula, Syria,[21] and Iraq; it became extinct around 1966.
- S. c. molybdophanes, Somali ostrich, southern Ethiopia, northeastern Kenya, and Somalia.[21] The neck and thighs are grey-blue, and during the mating season, the male's neck and thighs become brighter and bluer. The females are more brown than those of other subspecies.[24] It generally lives in pairs or alone, rather than in flocks. Its range overlaps with S. c. massaicus in northeastern Kenya.[24]
The population from Río de Oro was once separated as Struthio camelus spatzi because its eggshell pores were shaped like a teardrop and not round. However, as there is considerable variation of this character and there were no other differences between these birds and adjacent populations of S. c. camelus, the separation is no longer considered valid.[26] This population disappeared in the latter half of the 20th century. There were 19th-century reports of the existence of small ostriches in North Africa; these are referred to as Levaillant's ostrich (Struthio bidactylus) but remain a hypothetical form not supported by material evidence.[27]
Evolution
The earliest fossil of ostrich-like birds is the Palaeotis living near the Asiatic steppes[5] from the Middle Eocene, a mid-sized flightless bird that was originally believed to be a bustard. Apart from this enigmatic bird, the fossil record of the ostriches continues with several species of the modern genus Struthio which are known from the Early Miocene onwards. While the relationship of the African species is comparatively straightforward, a large number of Asian species of ostrich have been described from fragmentary remains, and their interrelationships and how they relate to the African ostriches are confusing. In China, ostriches are known to have become extinct only around or even after the end of the last ice age; images of ostriches have been found there on prehistoric pottery and petroglyphs.[28]
Several of these fossil forms are ichnotaxa (that is, classified according to the organism's footprints or other trace rather than its body) and their association with those described from distinctive bones is contentious and in need of revision pending more good material.[29]
In Subsaharan Africa:
- Struthio coppensi (Early Miocene of Elizabethfeld, Namibia)
- Struthio karingarabensis (Late Miocene – Early Pliocene of SW and CE Africa) – oospecies(?)
- Struthio kakesiensis (Laetolil Early Pliocene of Laetoli, Tanzania) – oospecies
- Struthio oldawayi (Early Pleistocene of Tanzania) – probably subspecies of S. camelus
- Struthio daberasensis (Early – Middle Pliocene of Namibia) – oospecies
- Struthio linxiaensis (Liushu Late Miocene of Yangwapuzijifang, China)
- Struthio orlovi (Late Miocene of Moldavia)
- Struthio karatheodoris (Late Miocene of Greece and Bulgaria)
- Struthio wimani (Early Pliocene of China and Mongolia)
- Struthio brachydactylus (Pliocene of Ukraine)
- Struthio chersonensis (Pliocene of SE Europe to WC Asia) – oospecies
- Asian Ostrich, Struthio asiaticus (Early Pliocene – Late Pleistocene of Central Asia to China ?and Morocco)
- Giant Ostrich, Struthio dmanisensis (Late Pliocene/Early Pleistocene of Dmanisi, Georgia)
- Struthio anderssoni (Pleistocene to Holocene in N China-Mongolia, to 8900 years BP [30]) [oospecies (?)]
Distribution and habitat
Ostriches formerly occupied Africa north and south of the Sahara, East Africa, Africa south of the rain forest belt, and much of Asia Minor.[5] Today ostriches prefer open land and are native to the savannas and Sahel of Africa, both north and south of the equatorial forest zone.[13] In Southwest Africa they inhabit the semi-desert or true desert. They rarely go above 100 m (330 ft).[5] Farmed ostriches in Australia have established feral populations.[1] The Arabian ostriches in the Near and Middle East were hunted to extinction by the middle of the 20th century. Ostriches have occasionally been seen inhabiting islands on the Dahlak Archipelago, in the Red Sea near Eritrea.Behaviour and ecology

Pair "dancing".

With their acute eyesight and hearing, ostriches can sense predators such as lions from far away. When being pursued by a predator, they have been known to reach speeds in excess of 70 km/h (43 mph),[5] and can maintain a steady speed of 50 km/h (31 mph), which makes the ostrich the world's fastest two-legged animal.[32][33] When lying down and hiding from predators, the birds lay their heads and necks flat on the ground, making them appear like a mound of earth from a distance, aided by the heat haze in their hot, dry habitat.[34][35]
When threatened, ostriches run away, but they can cause serious injury and death with kicks from their powerful legs.[13] Their legs can only kick forward.[36] Contrary to popular belief, ostriches do not bury their heads in sand to avoid danger.[37] This myth likely began with Pliny the Elder (AD 23–79), who wrote that ostriches "imagine, when they have thrust their head and neck into a bush, that the whole of their body is concealed."[38] This may have been a misunderstanding of their sticking their heads in the sand to swallow sand and pebbles,[39] or, as National Geographic suggests, of the defensive behavior of lying low, so that they may appear from a distance to have their head buried.[40]
Feeding
They mainly feed on seeds, shrubs, grass, fruit and flowers;[5][8] occasionally they also eat insects such as locusts. Lacking teeth, they swallow pebbles that act as gastroliths to grind food in the gizzard. When eating, they will fill their gullet with food, which is in turn passed down their esophagus in the form of a ball called a bolus. The bolus may be as much as 210 ml (7.1 US fl oz). After passing through the neck (there is no crop) the food enters the gizzard and is worked on by the aforementioned pebbles. The gizzard can hold as much as 1,300 g (46 oz), of which up to 45% may be sand and pebbles.[8] Ostriches can go without drinking for several days, using metabolic water and moisture in ingested plants,[41] but they enjoy liquid water and frequently take baths where it is available.[13] They can survive losing up to 25% of their body weight through dehydration.[42]Mating
Ostriches become sexually mature when they are 2 to 4 years old; females mature about six months earlier than males. As with other birds, an individual may reproduce several times over its lifetime. The mating season begins in March or April and ends sometime before September. The mating process differs in different geographical regions. Territorial males typically boom in defence of their territory and harem of two to seven hens;[43] the successful male may then mate with several females in the area, but will only form a pair bond with a 'major' female.[43]The cock performs with his wings, alternating wing beats, until he attracts a mate. They will go to the mating area and he will maintain privacy by driving away all intruders. They graze until their behaviour is synchronized, then the feeding becomes secondary and the process takes on a ritualistic appearance. The cock will then excitedly flap alternate wings again, and start poking on the ground with his bill. He will then violently flap his wings to symbolically clear out a nest in the soil. Then, while the hen runs a circle around him with lowered wings, he will wind his head in a spiral motion. She will drop to the ground and he will mount for copulation.[5] Ostriches raised entirely by humans may direct their courtship behaviour not at other ostriches, but toward their human keepers.[44]
-
 Only 15% of the surviving chicks reach 1 year of age
Only 15% of the surviving chicks reach 1 year of age
-
 Ostrich chick
Ostrich chick
-
 Ostrich chick and egg
Ostrich chick and egg
-
 Ostrich hen with chicks
Ostrich hen with chicks
-
 A female incubating eggs in a shallow nest on the ground in the Serengeti, Tanzania
A female incubating eggs in a shallow nest on the ground in the Serengeti, Tanzania
-
 Ostrich nest
Ostrich nest





Predators
As a flightless species in the rich biozone of the African savanna, the ostrich must face a variety of formidable predators throughout its life cycle. Animals that prey on ostriches of all ages may include cheetahs, lions, leopards, African hunting dogs, and spotted hyena.[5] Ostriches can often outrun most of their predators in a pursuit, so most predators will try to ambush an unsuspecting bird using obstructing vegetation or other objects.[49] A notable exception is the cheetah, which is the most prolific predator of adult ostriches due to its own great running speeds.[50]Common predators of nests and young ostriches include jackals, various birds of prey, warthogs, mongoose and Egyptian vultures.[43][51] If the nest or young are threatened, either or both of the parents may create a distraction, feigning injury.[8] However, they may sometimes fiercely fight predators, especially when chicks are being defended, and have been capable of killing even their largest enemies, the lions, in such confrontations.[40]
Physiology
Respiration
Anatomy

Diagrammatic location of the air sacs of the ostrich
Function
The ostrich is a thermoregulator and maintains a body temperature of 38.1–39.7 °C in its extreme living temperature conditions, such as the heat of the savanna and desert regions of Africa.[55] The ostrich utilizes its respiratory system via a costal pump for ventilation rather than a diaphragmatic pump as seen in most mammals.[53] Thus, they are able to use a series of air sacs connected to the lungs. The use of air sacs forms the basis for the three main avian respiratory characteristics:- Air is able to flow continuously in one direction through the lung, making it more efficient than the mammalian lung.
- It provides birds with a large residual volume, allowing them to breathe much more slowly and deeply than a mammal of the same body mass.
- It provides a large source of air that is used not only for gaseous exchange, but also for the transfer of heat by evaporation.[53]
To compensate for the large “dead” space, the ostrich trachea lacks valves to allow faster inspiratory air flow.[57] In addition, the total lung capacity of the respiratory system, (including the lungs and ten air sacs) of a 100 kg ostrich is about 15 L, with a tidal volume ranging from 1.2 to 1.5 L.[54][57] The tidal volume is seen to double resulting in a 16-fold increase in ventilation.[53] Overall, ostrich respiration can be thought of as a high velocity-low pressure system.[54] At rest, there is small pressure differences between the ostrich air sacs and the atmosphere, suggesting simultaneous filling and emptying of the air sacs.[57]
The increase in respiration rate from the low range to the high range is sudden and occurs in response to hyperthermia. Birds lack sweat glands, so when placed under stress due to heat, they heavily rely upon increased evaporation from the respiratory system for heat transfer. This rise in respiration rate however is not necessarily associated with a greater rate of oxygen consumption.[53] Therefore, unlike other birds, the ostrich is able to dissipate heat through panting without experiencing respiratory alkalosis by modifying ventilation of the respiratory medium. During hyperpnea ostriches pant at a respiratory rate of 40–60 cycles per minute, versus their resting rate of 6–12 cycles per minute.[54] Hot, dry and moisture lacking properties of the ostrich respiratory medium affects oxygen's diffusion rate (Henry's Law).[56]
Ostriches develop via Intussusceptive angiogenesis, a mechanism of blood vessel formation, characterizing many organs.[52] It is not only involved in vasculature expansion, but also in angioadaptation[58] of vessels to meet physiological requirements.[52] The use of such mechanisms demonstrates an increase in the later stages of lung development, along with elaborate parabronchial vasculature, and reorientation of the gas exchange blood capillaries to establish the crosscurrent system at the blood-gas barrier.[52] The blood–gas barrier (BGB) of their lung tissue is thick. The advantage of this thick barrier may be protection from damage by large volumes of blood flow in times of activity, such as running,[59] since air is pumped by the air sacs rather than the lung itself. As a result the capillaries in the parabronchi have thinner walls, permitting more efficient gaseous exchange.[53] In combination with separate pulmonary and systemic circulatory systems, it helps to reduce stress on the BGB.[52]
Circulation
Heart anatomy
The ostrich heart is a closed system, contractile chamber. It is composed of myogenic muscular tissue associated with heart contraction features. There is a double circulatory plan in place possessing both a pulmonary circuit and systemic circuit.[56]The ostrich’s heart has similar features to other avian species like having a conically shaped heart, and being enclosed by a pericardium layer.[60] Moreover, similarities also include a larger right atrium volume, and a thicker left ventricle to fulfil the systemic circuit.[60] The ostrich heart has three features that are absent in related birds:
- The right atrioventricular valve is fixed to the interventricular septum, by a thick muscular stock, which prevents back-flow of blood into the atrium when ventricular systole is occurring.[60] In the fowl this valve is only connected by a short septal attachment.[60]
- Pulmonary veins attach to the left atrium separately, and also the opening to the pulmonary veins are separated by a septum.[60]
- moderator bands, full of purkinje fibers, are found in different locations in the left and right ventricles.[60] These bands are associated with contractions of the heart and suggests this difference causes the left ventricle to contract harder to create more pressure for a completed circulation of blood around the body.[60]
The coronary arteries start in the right and left aortic sinus and provide blood to the heart muscle in a similar fashion to most other vertebrates.[62] Other domestic birds capable of flight have three or more coronary arteries that supply blood to the heart muscle. The blood supply by the coronary arteries are fashioned starting as a large branch over the surface of the heart. It then moves along the coronary groove and continues on into the tissue as interventricular branches toward the apex of the heart. The atria, ventricles, and septum are supplied of blood by this modality. The deep branches of the coronary arteries found with in the heart tissue are small and supply the interventricular and right atrioventricular valve with blood nutrients for which to carry out their processes. The interatrial artery of the ostrich is small in size and exclusively supplies blood to only part of the left auricle and interatrial septum.[26]
These purkinje fibers (p-fibers) found in the hearts moderator bands are a specialized cardiac muscle fiber that causes the heart to contract.[63] The purkinje cells are mostly found within both the endocardium and the sub-endocardium.[63] The sinoatrial node shows a small concentration of purkinje fibers, however, continuing through the conducting pathway of the heart the bundle of his shows the highest amount of these purkinje fibers.[63]
Blood composition
The red blood cell count per unit volume in the ostrich is about 40% of that of a human; however, the red blood cells of the ostrich are about three times larger than the red blood cells of a human.[64] The blood oxygen affinity, known as P50, is higher than that of both humans and similar avian species.[64] The reason for this decreased oxygen affinity is due to the hemoglobin configuration found in the ostrich blood.[64] The ostrich’s tetramer is composed of hemoglobin type A and D, compared to typical mammalian tetramers composed of hemoglobin type A and B; hemoglobin D configuration causes a decreased oxygen affinity at the site of the respiratory surface.[64]During the embryonic stage Hemoglobin E is present.[65] This subtype increases oxygen affinity in order to transport oxygen across the allantoic membrane of the embryo.[65] This can be attributed to the high metabolic need of the developing embryo, thus high oxygen affinity serves to satisfy this demand. When the chick hatches hemoglobin E diminishes while hemoglobin A and D increase in concentration.[65] This shift in hemoglobin concentration results in both decreased oxygen affinity and increased P50 value.[65]
Furthermore, the P50 value is influenced by differing organic modulators.[65] In the typical mammalian RBC 2,3 – DPG causes a lower affinity for oxygen. 2,3- DPG constitutes approximately 42–47%, of the cells phosphate of the embryonic ostrich.[65] However, the adult ostrich have no traceable 2,3- DPG.In place of 2,3-DPG the ostrich uses inositol polyphosphates (IPP), which vary from 1–6 phosphates per molecule.[65] In relation to the IPP, the ostrich also uses ATP to lower oxygen affinity.[65] ATP has a consistent concentration of phosphate in the cell.[65] Around 31% at incubation periods, and dropping to 16–20% in 36 day old chicks.[65] However, IPP has low concentrations, around 4%, of total phosphate concentration in embryonic stages; However, the IPP concentration jumps to 60% of total phosphate of the cell.[65] The majority of phosphate concentration switches from 2,3- DPG to IPP, suggesting the result of the overall low oxygen affinity is due to these varying polyphosphates.[65]
Concerning immunological adaptation, it was discovered that wild ostrich's have a pronounced non specific immunity defense, with blood content reflecting high values of lysosome, and phagocyte cells in medium. This is in contrast to domesticated ostriches, who in captivity develop high concentration of immunoglobulin antibodies in their circulation, indicating an acquired immunological response. It is suggested that this immunological adaptability may allow this species to have a high success rate of survival in variable environmental settings.[66]
Osmoregulation
Physiological challenges
The ostrich is an xeric animal, due to the fact that it lives in habitats that are both dry and hot.[56] Water is scarce in dry and hot environment, and this poses a challenge to the ostrich's water consumption. Also the ostrich is a ground bird and cannot fly to find water sources, which poses a further challenge. Because of their size, ostriches cannot easily escape the heat of their environment; however, they dehydrate less than their small bird counterparts because of their large surface area to volume ratio.[67] Hot, arid habitats pose osmotic stress, such as dehydration, which triggers the ostrich’s homeostatic response to osmoregulate.System overview
The ostrich is well adapted to hot, arid environments through specialization of excretory organs. The ostrich has an extremely long and developed colon the length of approximately 11-13m between the coprodeum and the paired caeca, which are around 80 cm long.[67] A well developed caeca is also found and in combination with the rectum forms the microbial fermentation chambers used for carbohydrate breakdown.[67] The catabolism of carbohydrates produces ~ 0.56 g of water that can be used internally.[56] The majority of their urine is stored in the coprodeum, and the faeces are separately stored in the terminal colon.[67] The coprodeum is located ventral to the terminal rectum and urodeum (where the ureters open).[53] Found between the terminal rectum and coprodeum is a strong sphincter.[53] The coprodeum and cloaca are the main osmoregulatory mechanisms used for the regulation and reabsorption of ions and water, or net water conservation.[53] As expected in a species inhabiting arid regions, dehydration causes a reduction in faecal water, or dry feces.[53] This reduction is believed to be caused by high levels of plasma aldosterone, which leads to rectal absorption of sodium and water.[53] Also expected is the production of hyperosmotic urine; cloacal urine has been found to be 800 mosmol/L.[53] The U:P (urine:plasma) ratio of the ostrich is therefore greater than one. Diffusion of water to the coprodeum (where urine is stored) from plasma across the epithelium is voided.[53] This void is believed to be caused by the thick mucosal layering of the coprodeum.[53]Ostriches have two kidneys, which are chocolate brown in color, granular in texture, and lie in a depression in the pelvic cavity of the dorsal wall.[68] They are covered by peritoneum and a layer of fat.[53] Each kidney is about 300 mm long, 70 mm wide, and divided into a cranial, middle, and caudal sections by large veins.[53] The caudal section is the largest, extends into the middle of the pelvis.[53] The ureters leave the ventral caudomedial surface and continue caudally, near the midline into the opening of the urodeum of the cloaca.[53] Although there is no bladder, a dilated pouch of ureter stores the urine until it is secreted continuously down from the ureters to the urodeum until discharged.[68]
Kidney function
Ostrich kidneys are fairly large, and so are able to hold significant amounts of solutes. Hence, ostriches drink relatively large volumes of water daily, and excrete generous quantities of highly concentrated urine. It is when drinking water is unavailable or withdrawn, that the urine becomes highly concentrated with uric acid and urates.[53] It seems that ostriches who normally drink relatively large amounts of water tend to rely on renal conservation of water when drinking water is scarce within the kidney system. Though there have been no official detailed renal studies conducted[69] on the flow rate (Poiseuille's Law) and composition of the ureteral urine in the ostrich, knowledge of renal function has been based on samples of cloacal urine, and samples or quantitative collections of voided urine.[53] Studies have shown that the amount of water intake, and dehydration impacts the plasma osmolality and urine osmolality within various sized ostriches. During a normal hydration state of the kidneys, young ostriches tend to have a measured plasma osmolality of 284 mOsm, and urine osmolality of 62 mOsm. Adults have higher rates with a plasma osmolality of 330 mOsm, and a urine osmolality of 163 mOsm. The osmolality of both plasma and urine can alter in regards to whether there is an excess or depleted amount of water present within the kidneys. An interesting fact of ostriches is that when water is freely available, the urine osmolality can reduce to 60–70 mOsm, not losing any necessary solutes from the kidneys when excess water is excreted.[53] Dehydrated or salt-loaded ostriches can reach a maximal urine osmolality of approximately 800 mOsm. When the plasma osmolality has been measured simultaneously with the maximal osmotic urine, it is seen that the urine:plasma ratio is 2.6:1, the highest encountered among avian species.[53] Along with dehydration, there is also a reduction in flow rate from 20 L·d-1 to only 0.3–0.5 L·d-1.In mammals and ostriches, the increase of the glomerular filtration rate (GFR) and urine flow rate (UFR) is due to a high protein diets. As seen in various studies, scientists have measured clearance of creatinine, a fairly reliable marker of glomerular filtration rate (GFR).[53] It has been seen that during normal hydration within the kidneys, the glomerular filtration rate is approximately 92 ml/min. However, when an ostrich experiences dehydration for at least 48 hours (2 days), this value diminishes to only 25% of the hydrated GFR rate. Thus in response to the dehydration, ostrich kidneys secrete small amounts of very viscous glomerular filtrates that have not been broken down, and return them to the circulatory system through blood vessels. The reduction of GFR during dehydration is extremely high, and so the fractional excretion of water (urine flow rate as a percentage of GFR) drops down from 15% at normal hydration to 1% during dehydration.[53]
Water intake and turnover
Ostriches employ adaptive features to manage the dry heat and solar radiation in their habitat. Ostriches will drink available water; however, they are limited in accessing water by being flightless. They are also able to harvest water through dietary means, consuming plants such as the Euphorbia heterochroma that hold up to 87% water.[53]Water mass accounts for 68% of body mass in adult ostriches; this is down from 84% water mass in 35-day-old chicks. The differing degrees of water retention are thought to be a result of varying body fat mass.[53] In comparison to smaller birds ostriches have a lower evaporative water loss resulting from their small body surface area per unit weight.[56]
When heat stress is at its maximum, ostriches are able to recover evaporative loss by using a metabolic water mechanism to counter the loss by urine, feces, and respiratory evaporation. An experiment to determine the primary source of water intake in the ostrich indicated that while the ostrich does employ a metabolic water production mechanism as a source of hydration, the most important source of water is food. When ostriches were restricted to the no food or water condition, the metabolic water production was only 0.5 L·d-1, while total water lost to urine, feces and evaporation was 2.3 L·d-1. When the birds were given both water and food, total water gain was 8.5 L·d-1. In the food only condition total water gain was 10.1 L·d-1. These results show that the metabolic water mechanism is not able to sustain water loss independently, and that food intake, specifically of plants with a high water content such as Euphorbia heterochroma, is necessary to overcome water loss challenges in the ostrich's arid habitat.[53]
In times of water deprivation, urine electrolyte and osmotic concentration increases while urination rate decreases. Under these conditions urine solute:plasma ratio is approximately 2.5, or hyperosmotic; that is to say that the ratio of solutes to water in the plasma is shifted down whereby reducing osmotic pressure in the plasma. Water is then able to be held back from excretion, keeping the ostrich hydrated, while the passed urine contains higher concentrations of solute. This mechanism exemplifies how renal function facilitates water retention during periods of dehydration stress.[56][70]
Nasal glands
A number of avian species use nasal salt glands, alongside their kidneys, to control hypertonicity in their blood plasma.[71] However, the ostrich shows no nasal glandular function in regard to this homeostatic process.[71] Even in a state of dehydration, which increases the osmolality of the blood, nasal salt glands show no sizeable contribution of salt elimination.[71] Also, the overall mass of the glands was less than that of the duck’s nasal gland.[71] The ostrich, having a heavier body weight, should have larger, heavier nasal glands to more effectively excrete salt from a larger volume of blood, but this is not the case. These unequal proportions contribute to the assumption that the ostrich’s nasal glands do not play any role in salt excretion. The nasal glands may be the result of an ancestral trait, which is no longer needed by the ostrich, but has not been bred out of their gene pool.[citation needed]Biochemistry
The majority of the ostrich’s internal solutes are made up of sodium ions (Na+), potassium ions (K+), chloride ions (Cl-), total short-chain fatty acids (SCFA), and acetate.[67] The caecum contains a high water concentration with reduced levels nearing the terminal colon, and exhibits a rapid fall in Na+ concentrations and small changes in K+ and Cl-.[67] The colon is divided into three sections and take part in solute absorption. The upper colon largely absorbs Na+ and SCFA, and partially absorbs KCl.[67] The middle colon absorbs Na+, SCFA, with little net transfer of K+ and Cl-.[67] The lower colon then slightly absorbs Na+ and water, and secretes K+. There is no net movements of Cl- and SCFA found in the lower colon.[67]When the ostrich is in a dehydrated state plasma osmolality, Na+, K+, and Cl- ions all increase, however, K+ ions returned to controlled concentration.[72] The ostrich also experiences an increase in haematocrit, resulting in a hypovolemic state.[72] Two antidiuretic hormones, Arginine vasotocin (AVT) and angiotensin (AII) are increased in blood plasma as a response to hyperosmolality and hypovolemia.[72] AVT triggers antidiuretic hormone (ADH) which targets the nephrons of the kidney.[56] ADH causes a reabsorption of water from the lumen of the nephron to the extracellular fluid osmotically.[56] These extracellular fluids then drain into blood vessels, causing a rehydrating effect.[56] This drainage prevents loss of water by both lowering volume and increasing concentration of the urine.[56] Angiotensin, on the other hand, causes vasoconstriction on the systemic arterioles, and acts as a dipsogen for ostriches.[56] Both of these antidiuretic hormones work together to maintain water levels in the body that would normally be lost due to the osmotic stress of the arid environment.
The end-product of catabolism of protein metabolism in animals is nitrogen.[56] Animals must excrete this in the form of nitrogenous compounds.[56] Ostriches are uricotelic. They excrete nitrogen as the complex nitrogenous waste compound uric acid, and related derivatives.[56] Uric acid's low solubility in water gives a semi-solid paste consistency to the ostrich's nitrogenous waste.[56]
Thermoregulation
Ostriches are homeothermic endotherms; they regulate a constant body temperature via regulating their metabolic heat rate.[56] They closely regulate their core body temperature, but their appendages may be cooler in comparison as found with regulating species.[56] The temperature of their beak, neck surfaces, lower legs, feet and toes are regulated through heat exchange with the environment.[73] Up to 40% of their produced metabolic heat is dissipated across these structures, which account for about 12% of their total surface area.[73] Total evaporative water loss (TEWL) is statistically lower in the ostrich than in membering ratites.[74]As ambient temperature increases, dry heat loss decreases, but evaporative heat loss increases because of increased respiration.[73] As ostriches experience high ambient temperatures, ~50 °C, they become slightly hyperthermic; however, they can maintain a stable body temperature, ~40 °C, for up to 8 hours in these conditions.[75] When dehydrated, the ostrich minimises water loss, causing the body temperature to increase further.[75] When the body heat is allowed to increase the temperature gradient between the ostrich and ambient heat is equilibrated.[56]
Physical adaptations
Ostriches have developed a comprehensive set of behavioural adaptations for thermoregulation, such as altering their feathers.[53] Ostriches display a feather fluffing behaviour that aids them in thermoregulation by regulating convective heat loss at high ambient temperatures.[73] They may also physically seek out shade in times of high ambient temperatures. When feather fluffing, they contract their muscles to raise their feathers to increase the air space next to their skin.[56] This air space provides an insulating thickness of 7 cm.[76] The ostrich will also expose the thermal windows of their unfeathered skin to enhance convective and radiative loss in times of heat stress.[74] At higher ambient temperatures lower appendage temperature increases to 5 °C difference from ambient temperature.[73] Neck surfaces are around 6–7 °C difference at most ambient temperatures, except when temperatures are around 25 °C, it was only 4 °C above ambient.[73]At low ambient temperatures the ostrich utilizes feather flattening, which conserves body heat through insulation. The low conductance coefficient of air allows less heat to be lost to the environment.[56] This flattening behavior compensate for ostrich's rather poor cutaneous evaporative water loss (CEWL).[77] These feather heavy areas such as the body, thighs and wings do not usually vary much from ambient temperatures due to this behavioural controls.[73] This ostrich will also cover its legs to reduce heat loss to the environment, along with undergoing piloerection and shivering when faced with low ambient temperatures.
Internal adaptations
The use of countercurrent heat exchange with blood flow allows for regulated conservation/ elimination of heat of appendages.[56] When ambient temperatures are low, heterotherms will constrict their arterioles to reduce heat loss along skin surfaces.[56] The reverse occurs at high ambient temperatures, arterioles dilate to increase heat loss.[56]At ambient temperatures below their body temperatures (thermal neutral zone (TNZ)), ostriches decrease body surface temperatures so that heat loss occurs only across about 10% of total surface area.[73] This 10% include critical areas that require blood flow to remain high to prevent freezing, such as their eyes.[73] Their eyes and ears tend to be the warmest regions.[73] It has been found that temperatures of lower appendages were no more than 2.5 °C above ambient temperature, which minimizes heat exchange between feet, toes, wings, and legs.[73]
Both the Gular and air sacs, being close to body temperature, are the main contributors to heat and water loss.[75] Surface temperature can be affected by the rate of blood flow to a certain area, and also by the surface area of the surrounding tissue.[56] The ostrich reduces blood flow to the trachea to cool itself, and vasodilates its blood vessels around the gular region to raise the temperature of the tissue.[75] The air sacs are poorly vascularized but show an increased temperature, which aids in heat loss.[75]
Ostriches have evolved a 'selective brain cooling' mechanism as a means of thermoregulation. This modality allows the ostrich to manage the temperature of the blood going to the brain in response to the extreme ambient temperature of the surroundings. The morphology for heat exchange occurs via cerebral arteries and the ophthalmic rete, a network of arteries originating from the ophthalmic artery. The ophthalmic rete is analogous to the carotid rete found in mammals, as it also facilitates transfer of heat from arterial blood coming from the core to venous blood returning from the evaporative surfaces at the head.
[78]
Researchers suggest that ostriches also employ a ‘selective brain warming’ mechanism in response to cooler surrounding temperatures in the evenings. The brain was found to maintain a warmer temperature when compared to carotid arterial blood supply. Researchers hypothesize three mechanisms for this finding. They first suggest a possible increase in metabolic heat production within the brain tissue itself to compensate for the colder arterial blood arriving from the core. They also speculate that there is an overall decrease in cerebral blood flow to the brain. Finally, they suggest that warm venous blood perfusion at the ophthalmic rete facilitates warming of cerebral blood that supplies the hypothalamus. Further research will need to be done to find how this occurs.[78]
Breathing adaptations
The ostrich has no sweat glands, and under heat stress they rely on panting to reduce their body temperature.[75] Panting increases evaporative heat (and water) loss from its respiratory surfaces, therefore forcing air and heat removal without the loss of metabolic salts.[74] Panting allows the ostrich to have a very effective respiratory evaporative water loss (REWL). Heat dissipated by respiratory evaporation increases linearly with ambient temperature, matching the rate of heat production.[53] As a result of panting the ostrich should eventually experience alkalosis.[56] However, The CO2 concentration in the blood does not change when hot ambient temperatures are experienced.[75] This effect is caused by a lung surface shunt.[75] The lung is not completely shunted, allowing enough oxygen to fulfill the bird’s metabolic needs.[75] The ostrich utilizes gular fluttering, rapid rhythmic contraction and relaxation of throat muscles, in a similar way to panting.[56] Both these behaviors allow the ostrich to actively increase the rate of evaporative cooling.[56]In hot temperatures water is lost via respiration.[56] Moreover, varying surface temperatures within the respiratory tract contribute differently to overall heat and water loss through panting.[75] The surface temperature of the gular area is 38 °C; that of the tracheal area, between 34 °C and 36 °C; and that of both anterior and posterior air sacs, 38 °C.[75] The long trachea, being cooler than body temperature, is a site of water evaporation.[75]
As ambient air becomes hotter, additional evaporation can take place lower in the trachea making its way to the posterior sacs, shunting the lung surface.[75] The trachea acts as a buffer for evaporation because of the length, and the controlled vascularization.[75] The Gular is also heavily vascularized; its purpose is for cooling blood, but also evaporation, as previously stated. Air flowing through the trachea can be either laminar or turbulent depending on the state of the bird.[56] When the ostrich is breathing normally, under no heat stress, air flow is laminar.[75] When the ostrich is experiencing heat stress from the environment the air flow is considered turbulent.[75] This suggests that laminar air flow causes little to no heat transfer, while under heat stress turbulent airflow can cause maximum heat transfer within the trachea.[75]
Development
Ostrich embryos during development transition from ectotherms to endotherms. This change requires an increase in energy production or ATP. Research has found that increased phosphorylation of AMP activated protein kinase is part of a positive feedback mechanism influencing an increase of transcription factor concentrations that facilitate an increase of mitochondrial biosynthesis. Part of the mechanism behind the increase in mitochondrial abundance has been described here and is a key factor in transitioning from an ectotherm to an endotherm for these developing chicks.[79]
Metabolism
Ostriches are able to attain their necessary energetic requirements via the oxidation of absorbed nutrients. Much of the metabolic rate in animals is dependent upon their allometry, the relationship between body size to shape, anatomy, physiology and behaviour of an animal. Hence, it is plausible to state that metabolic rate in animals with larger masses is greater than animals with a smaller mass.When a bird is inactive, unfed, and the ambient temperature (i.e. in the thermo-neutral zone) is high, the energy expended is at its minimum. This level of expenditure is better known as the basal metabolic rate (BMR), and can be calculated by measuring the amount of oxygen consumed during various activities.[53] Therefore in ostriches we see use of more energy when compared to smaller birds in absolute terms, but less per unit mass.
A key point when looking at the ostrich metabolism is to note that it is a non-passerine bird. Thus, BMR in ostriches is particularly low with a value of only 0.113 ml O2 g−1 h−1. This value can further be described using Kleiber's law, which relates the BMR to the body mass of an animal.[80]
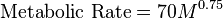
where
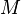 is body mass, and metabolic rate is measured in kcal per day.
is body mass, and metabolic rate is measured in kcal per day.In ostriches, a BMR (ml O2 g−1 h−1) = 389 kg0.73, describing a line parallel to the intercept with only about 60% in relation to other non-passerine birds.[53]
Along with BMR, energy is also needed for a range of other activities. If the ambient temperature is lower than the thermo-neutral zone, heat is produced to maintain body temperature.[53] So, the metabolic rate in a resting, unfed bird, that is producing heat is known as the standard metabolic rate (SMR) or resting metabolic rate(RMR). The ostrich SMR has been seen to be approximately 0.26 ml O2 g−1 h−1, almost 2.3 times the BMR.[53] On another note, animals that engage in extensive physical activity employ substantial amounts of energy for power. This is known as the maximum metabolic scope. In an ostrich, it is seen to be at least 28 times greater than the BMR. Likewise, the daily energy turnover rate for an ostrich with access to free water is 12,700 kJ·d−1, equivalent to 0.26 ml O2 g−1 h−1.[53]
Status and conservation
The wild ostrich population has declined drastically in the last 200 years, with most surviving birds in reserves or on farms.[5] However, its range remains very large (9,800,000 square kilometres (3,800,000 sq mi)), leading the IUCN and BirdLife International to treat it as a species of Least Concern.[1] Of its 5 subspecies, the Middle Eastern ostrich (S. c. syriacus) became extinct around 1966, and the North African ostrich (S. c. camelus) has declined to the point where it now is included on CITES Appendix I and some treat it as Critically Endangered.[22][23][24]
Scene with ostriches, Roman mosaic, 2nd century AD
Ostriches and humans
Ostriches have inspired cultures and civilizations for 5,000 years in Mesopotamia and Egypt. A statue of Arsinoe II of Egypt riding an ostrich was found in a tomb in Egypt.[81] Hunter-gatherers in the Kalahari use ostrich eggshells as water containers, punching a hole in them.[5][82] The presence of such eggshells with engraved hatched symbols dating from the Howiesons Poort period of the Middle Stone Age at Diepkloof Rock Shelter in South Africa suggests ostriches were an important part of human life as early as 60,000 BP.[83]Hunting and farming

Fashion accessories made from ostrich feathers, Amsterdam, 1919
Ostriches were farmed for their feathers in South Africa beginning in the 19th century. According to Frank G. Carpenter, the English are credited with first taming ostriches outside Cape Town. Farmers captured baby ostriches and raised them successfully on their property, and were able to obtain a crop of feathers every seven to eight months instead of killing wild ostriches for their feathers.[84]
It is claimed that ostriches produce the strongest commercial leather.[85] Ostrich meat tastes similar to lean beef and is low in fat and cholesterol, as well as high in calcium, protein and iron. Uncooked, it is dark red or cherry red, a little darker than beef.[86]
Attacks
Ostriches typically avoid humans in the wild, since they correctly assess humans as potential predators. If approached, they often run away, but sometimes ostriches can be very aggressive when threatened, especially if cornered, and may also attack if they feel the need to defend their territories or offspring. Similar behaviors are noted in captive or domesticated ostriches, which retain the same natural instincts and can occasionally respond aggressively to stress. When attacking a person, ostriches deliver slashing kicks with their powerful feet, armed with long claws, with which they can disembowel or kill a person with a single blow.[87] In one study of ostrich attacks, it was estimated that two to three attacks that result in serious injury or death occur each year in the area of Oudtshoorn, South Africa, where a large number of ostrich farms are set next to both feral and wild ostrich populations.[48]
A Jacksonville, Florida, man with an ostrich-drawn cart, circa 1911
Racing
See also: List of racing forms § Animal racing
In some countries, people race each other on the backs of ostriches. The practice is common in Africa[88] and is relatively unusual elsewhere.[89] The ostriches are ridden in the same way as horses with special saddles, reins, and bits. However, they are harder to manage than horses.[90]The racing is also a part of modern South African culture.[91] Within the United States, a tourist attraction in Jacksonville, Florida called 'The Ostrich Farm' opened up in 1892; it and its races became one of the most famous early attractions in the history of Florida.[92]
In the United States, Chandler, Arizona hosts the annual 'Ostrich Festival', which features ostrich races.[93][94] Racing has also occurred at many other locations such as Virginia City in Nevada, Canterbury Park in Minnesota,[95] Prairie Meadows in Iowa, Ellis Park in Kentucky,[96] and the Fairgrounds in New Orleans, LA.[97]
No comments:
Post a Comment
Note: Only a member of this blog may post a comment.